

The National Institute of Certified Floor-Covering Inspectors (NICFI) is a
non-profit association of independent floor covering inspectors. NICFI members are expert flooring failure analysts and expert witnesses in flooring problems and failures.
NICFI members are experienced in the inspection and failure analysis of
woven carpet, broadloom carpet, resilient tile, rubber tile, cork tile, ceramic tile, resilient sheet vinyl, vinyl composition tile, LVT, hardwood, laminate, solid vinyl,
and specialty flooring materials.
NICFI members attend the annual NICFI Convention in Dalton, Georgia, to increase their knowledge and update their skills. The Annual Convention includes updates on indicative field testing
procedures, tools, products and product defects. The NICFI is the only Non-Profit Association in the Flooring Industry with membership comprised solely of Certified Floor Covering Inspectors.



PHOTOS
IS THIS YOUR COMPLAINT?
































































































































































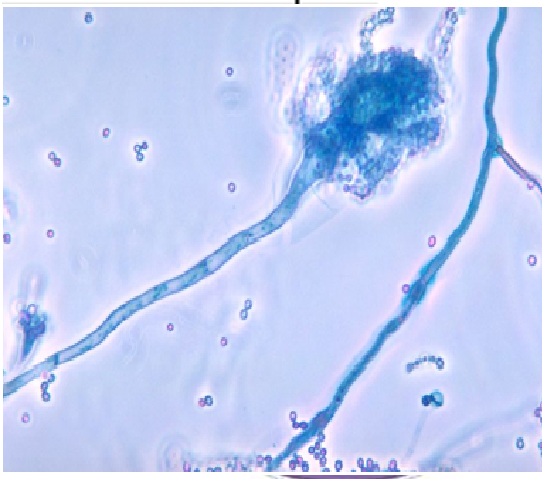
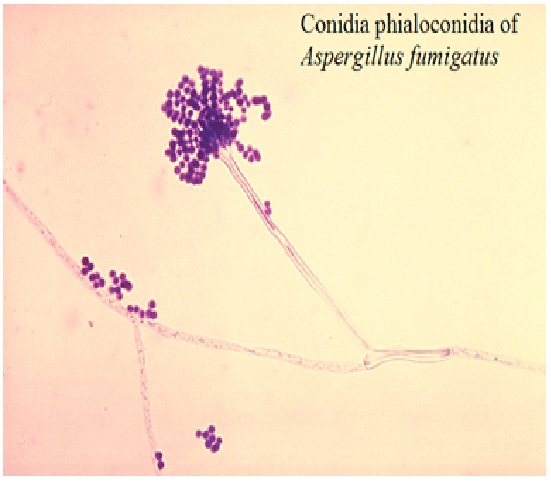
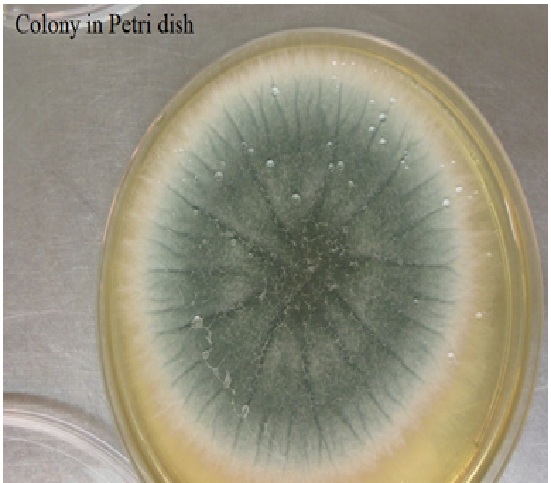
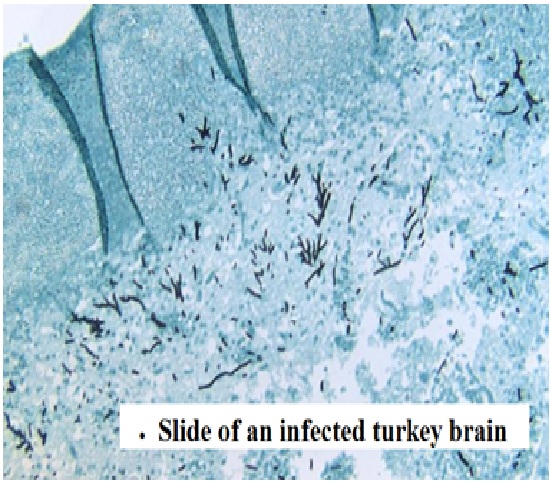
1. ASPERGILLUS FUMIGATUS FUNGUS
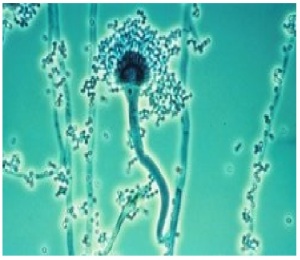
Aspergillus fumigatus is a fungus of the genus Aspergillus, and is one of the most common Aspergillus species to cause disease in individuals with an immunodeficiency.
A. fumigatus, a saprotroph widespread in nature, is typically found in soil and decaying organic matter, such as compost heaps, where it plays an essential role in carbon and nitrogen recycling. Colonies of the fungus produce from conidiophores thousands of minute grey-green conidia (2–3 μm) that readily become airborne. For many years, A. fumigatus was thought to only reproduce asexually, as neither mating nor meiosis had ever been observed. In 2008, however, A. fumigatus was shown to possess a fully functional sexual reproductive cycle, 145 years after its original description by Fresenius.
The fungus is capable of growth at 37 °C/99 °F (normal human body temperature), and can grow at temperatures up to 50 °C/122 °F, with conidia surviving at 70 °C/158 °F—conditions it regularly encounters in self-heating compost heaps. Its spores are ubiquitous in the atmosphere, and it is estimated that everybody inhales several hundred spores each day; typically these are quickly eliminated by the immune system in healthy individuals. In immunocompromised individuals, such as organ transplant recipients and people with AIDS or leukemia, the fungus is more likely to become pathogenic, over-running the host's weakened defenses and causing a range of diseases generally termed aspergillosis.
When the fermentation broth of A. fumigatus was screened, a number of indolic alkaloids with antimitotic properties were discovered. The compounds of interest have been of a class known as tryprostatins, with spirotryprostatin B being of special interest as an anticancer drug.
A. fumigatus grown on certain building materials can produce genotoxic and cytotoxic mycotoxins, such as gliotoxin.
A. fumigatus is the most frequent cause of invasive fungal infection in immunosuppressed individuals, which include patients receiving immunosuppressive therapy for autoimmune or neoplastic disease, organ transplant recipients, and AIDs patients. A. fumigatus primarily causes invasive infection in the lung and represents a major cause of morbidity and mortality in these individuals. Additionally, A. fumigatus can cause chronic pulmonary infections or allergic disease in immunocompetent hosts.
Innate Immune Response
Inhalational exposure to airborne conidia is continuous due to their ubiquitous distribution in the environment. However, in healthy individuals the innate immune system is an efficacious barrier to A. fumigatus infection. A large portion of inhaled conidia are cleared by the mucociliary action of the respiratory epithelium. Due to the small size of conidia, many conidia deposit in alveoli where they interact with epithelial and innate effector cells. Alveolar macrophages phagocytize and destroy conidia within their phagosomes. Epithelial cells, specifically type II pneumocytes, also internalize conidia which traffic to the lysosome where ingested conidia are destroyed. First line immune cells also serve to recruit neutrophils and other inflammatory cells through release of cytokines and chemokines induced by ligation of specific fungal motifs to pathogen recognition receptors. Neutrophils are essential for aspergillosis resistance, as demonstrated in neutropenic individuals, and are capable of sequestering both conidia and hyphae through distinct, non-phagocytic mechanisms. Hyphae are too large for cell-mediated internalization, and thus neutrophil-mediated NADPH-oxidase induced damage represents the dominant host defense against hyphae. In addition to these cell-mediated mechanisms of elimination, antimicrobial peptides secreted by the airway epithelium contribute to host defense.
Invasion
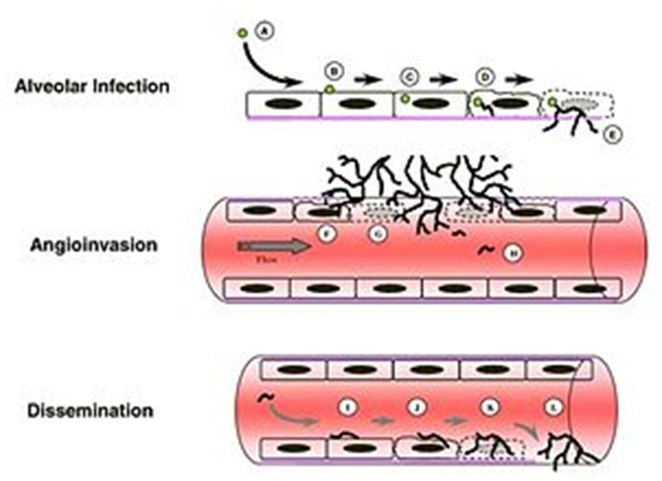
Schematic of Invasive Aspergillus Infection. Hyphae germinate either within an epithelial cell or within the alveoli. Hyphae extend through the epithelial cells, eventually invading and traversing endothelial cells of the vasculature. In rare cases, hyphal fragments break off and disseminate through the blood stream.
Immunosuppressed individuals are susceptible to invasive A. fumigatus infection, which most commonly manifests as invasive pulmonary aspergillosis. Inhaled conidia that evade host immune destruction are the progenitors of invasive disease. These conidia emerge from dormancy and make a morphological switch to hyphae by germinating in the warm, moist, nutrient-rich environment of the pulmonary alveoli. Germination occurs both extracellularly or in type II pneumocyte endosomes containing conidia. Following germination, filamentous hyphal growth results in epithelial penetration and subsequent penetration of the vascular endothelium. The process of angioinvasion causes endothelial damage and induces a proinflammatory response, tissue factor expression and activation of the coagulation cascade. This results in intravascular thrombosis and localized tissue infarction, however, dissemination of hyphal fragments is usually limited. Dissemination through the blood stream only occurs in severely immunocomprimised individuals.
Nutrient Acquisition
A. fumigatus must acquire nutrients from its external environment in order to survive and flourish within its host. Many of the genes involved in such processes have been shown to impact virulence through experiments involving genetic mutation. Examples of nutrient uptake include that of metals, nitrogen, and macromolecules such as peptides.
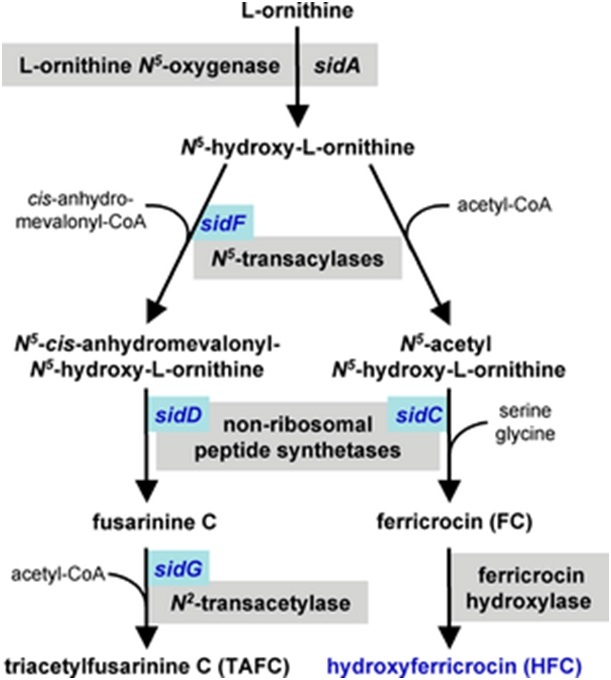
Proposed Siderophore Biosynthetic Pathway of Aspergillus fumigatus: sidA catalyzes the first step in the biosythesis of both the extracellular siderophore triacetylfusarinine C and intracellular ferricrocin
Iron Acquisition
Iron is a necessary cofactor for many enzymes, and can act as a catalyst in the electron transport system. A. fumigatus has two mechanisms for the uptake of iron, reductive iron acquisition and siderophore-mediated. Reductive iron acquisition includes conversion of iron from the ferric (Fe+3) to the ferrous (Fe+2) state and subsequent uptake via FtrA, an iron permease. Targeted mutation of the ftrA gene did not induce a decrease in virulence in the murine model of A. fumigatus invasion. In contrast, targeted mutation of sidA, the first gene in the sideophore biosynthesis pathway, proved sideophore-mediated iron uptake to be essential for virulence. Mutation of the downstream sideophore biosynthesis genes sidC, sidD, sidF and sidG resulted in strains of A. fumigatus with similar decreases in virulence. These mechanisms of iron uptake appear to work in parallel and both are upregulated in response to iron starvation.
Nitrogen assimilation
A. fumigatus can survive on a variety of different nitrogen sources, and the assimilation of nitrogen is of clinical importance as it has been shown to affect virulence. Proteins involved in nitrogen assimilation are transcriptionally regulated by the AfareA gene in A. fumigatus. Targeted mutation of the afareA gene showed a decrease in onset of mortality in a mouse model of invasion. The Ras[disambiguation needed] regulated protein RhbA has also been implicated in nitrogen assimilation. RhbA was found to be transcriptionally upregulated following contact of A. fumigatus with human endothelial cells, and strains with targeted mutation of the rhbA gene showed decreased growth on poor nitrogen sources and reduced virulence in vivo.
Proteinases
The human lung contains large quantities of collagen and elastin, proteins that allow for tissue flexibility. Aspergillus fumigatus produces and secretes elastases, proteases that cleave elastin in order to break down these macromolecular polymers for uptake. A significant correlation between the amount of elastase production and tissue invasion was first discovered in 1984. Clinical isolates have also been found to have greater elastase activity than environmental strains of A. fumigatus. A number of elastases have been characterized, including those from the serine protease, aspartic protease, and metalloprotease families. Yet, the large redundancy of these elastases has hindered the identification of specific effects on virulence.
Secondary metabolites in fungal development
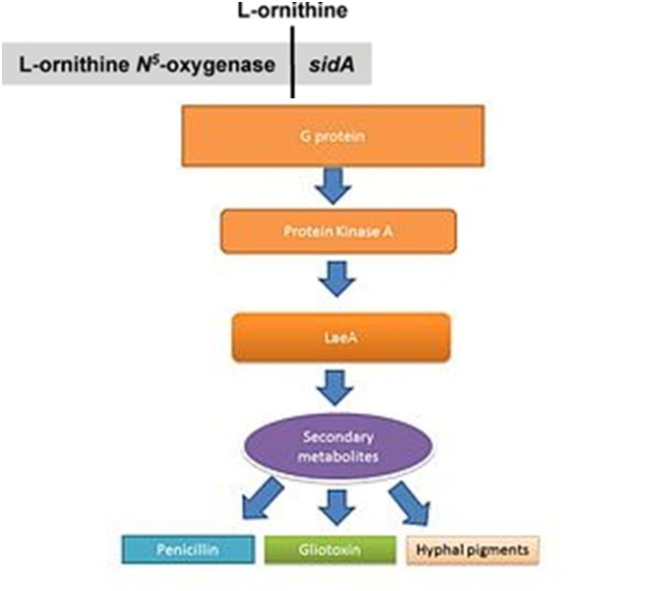
The transcription factor LaeA regulates the expression of several genes involved in secondary metabolite production in Aspergillus spp.
The life cycle of filamentous fungi including Aspergillus spp. consists of two phases: a hyphal growth phase and a reproductive (sporulation) phase. The switch between growth and reproductive phases of these fungi is regulated in part by the level of secondary metabolite production. It is believed the secondary metabolites are produced to activate sporulation and pigments required for sporulation structures. G protein signaling regulates secondary metabolite production. Genome sequencing has revealed 40 potential genes involved in secondary metabolite production including mycotoxins, which are produced at the time of sporulation.
Gliotoxin
Gliotoxin is a mycotoxin capable of altering host defenses through immunosuppression. Neutrophils are the principal targets of gliotoxin. Gliotoxin interrupts the function of leukocytes by inhibiting migration and superoxide production and causes apoptosis in macrophages. Gliotoxin disrupts the proinflammatory response through inhibition of NF-κB.
Transcriptional Regulation of Gliotoxin
LaeA and GliZ are transcription factors known to regulate the production of gliotoxin. LaeA is a universal regulator of secondary metabolite production in Aspergillus spp. LaeA influences the expression of 9.5% of the A. fumigatus genome, including many secondary metabolite biosynthesis genes such as nonribosomal peptide synthetases (NRPSs). The production of numerous secondary metabolites, including gliotoxin, were impaired in an LaeA mutant (ΔlaeA) strain. The ΔlaeA mutant showed increased susceptibility to macrophage phagocytosis and decreased ability to kill neutrophils ex vivo. LaeA regulated toxins, besides gliotoxin, likely have a role in virulence since loss of gliotoxin production alone did not recapitulate the hypo-virulent ∆laeA pathotype.